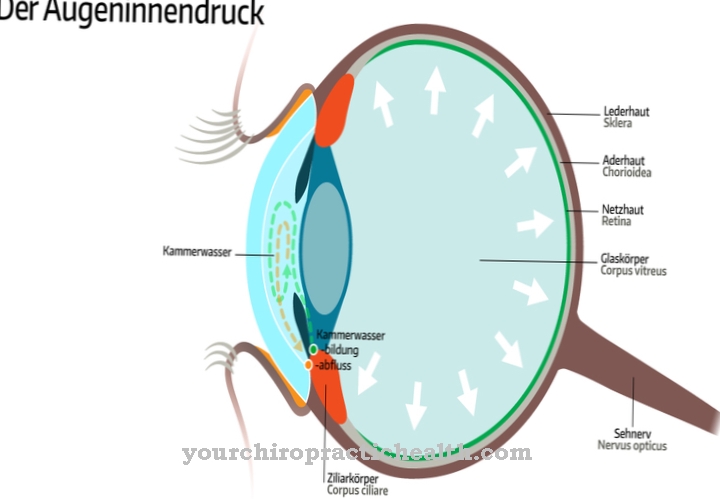
ال نهاية هي المرحلة النهائية في تكرار الحمض النووي. يسبقه البدء والاستطالة. يمكن أن يؤدي الإنهاء المبكر للنسخ المتماثل إلى ظهور بروتينات مختصرة وبالتالي حدوث طفرة.
ما هو الإنهاء؟

أثناء التكرار أو المضاعفة ، يتم مضاعفة الحمض النووي الناقل للمعلومات الجينية في الخلايا الفردية. تحدث الازدواجية وفقًا للمبدأ شبه المحافظ وعادة ما تؤدي إلى تكرار دقيق للمعلومات الجينية. يتم تشغيل النسخ المتماثل خلال مرحلة التوليف ، قبل مرحلة الانقسام الفتيلي ، وبالتالي يحدث قبل انقسام نواة الخلية.
في بداية النسخ المتماثل ، يتم فصل خيوط الحمض النووي المزدوجة إلى خيوط مفردة ، تتشكل عليها خيوط تكميلية جديدة. يتم تحديد كل خيط DNA من خلال التسلسل الأساسي للخيط المعاكس. يحدث تكرار الحمض النووي في عدة مراحل. الإنهاء هو المرحلة الثالثة وبالتالي الأخيرة من النسخ المتماثل. الإنهاء يسبقه البدء والاستطالة.
المصطلح المرادف للتعبير عن الإنهاء في هذا السياق هو التسمية مرحلة الإنهاء. يعني الإنهاء هنا "إجهاض" أو "إنهاء". أثناء الإنهاء ، ينفصل خيط الرنا المرسال المتشكل حديثًا عن الحمض النووي الفعلي. يقترب عمل بوليميراز الحمض النووي ببطء من نهايته. لا ينبغي الخلط بين إنهاء تكرار الحمض النووي مع إنهاء تكرار الحمض النووي الريبي.
الوظيفة والمهمة
في مرحلة التكرار من البدء ، يتم تنظيم النسخ المتماثل بشكل أساسي. يتم تحديد نقطة البداية للنسخ المتماثل ويحدث ما يسمى بالتمهيد. بعد البدء ، تبدأ البلمرة ، والتي تحدث خلالها مرحلة الاستطالة. يفصل إنزيم بوليميراز DNA الخيوط التكميلية للحمض النووي إلى خيوط مفردة ويقرأ قواعد الخيوط المفردة واحدة تلو الأخرى. في هذه المرحلة ، تحدث المضاعفة شبه المتقطعة ، والتي تتضمن مرحلة أخرى من التحضير.
فقط بعد البدء والاستطالة تتبع مرحلة الإنهاء ضمن النسخ المتماثل. يختلف الإنهاء من شكل الحياة إلى شكل الحياة. في حقيقيات النوى مثل البشر ، يتكون الحمض النووي في حلقة. يتضمن أيضًا متواليات الإنهاء التي تتوافق مع تسلسلين مختلفين ، كل منهما مناسب لشوكة النسخ المتماثل.
عادة لا يتم تشغيل الإنهاء بواسطة آليات خاصة. بمجرد تشغيل شوكتي نسخ معًا أو انتهاء DNA ، يتم إنهاء النسخ المتماثل تلقائيًا في هذه المرحلة. يتم إنهاء النسخ المتماثل في آلية تلقائية.
تسلسلات الإنهاء هي عناصر تحكم. إنها تضمن وصول مرحلة النسخ المتماثل إلى نقطة نهاية محددة بطريقة مضبوطة على الرغم من سرعات النسخ المختلفة في شوكات النسخ المتماثل. تتوافق جميع مواقع الإنهاء مع مواقع الربط لبروتين Tus ، "نهاية استخدام المادة". يمنع هذا البروتين الهيليكس التكاثري DnaB وبالتالي يوقف التكرار.
في حقيقيات النوى ، تظل خيوط الحلقة المنسوخة متصلة ببعضها البعض حتى بعد التكرار. الاتصال يتوافق مع نقطة المحطة. فقط بعد انقسام الخلية يتم فصلهم عن طريق عمليات مختلفة وبالتالي يمكن تقسيمهم. يبدو أن الاتصال الذي يبقى حتى بعد انقسام الخلية يلعب دورًا في التوزيع الخاضع للرقابة.
هناك آليتان رئيسيتان تلعبان دورًا في الفصل النهائي لحلقات الحمض النووي. وتشارك إنزيمات مثل النوع الأول والنوع الثاني توبويزوميراز في الفصل. أخيرًا ، يتعرف البروتين الإضافي على كودون الإيقاف أثناء الإنهاء. يؤدي هذا إلى سقوط البولي ببتيد من الريبوسوم ، حيث لا يتوفر t-RNA مع مضاد كودون مناسب لكودون الإيقاف. في النهاية ، ينقسم الريبوسوم إلى وحدتين فرعيتين.
الامراض والاعتلالات
جميع عمليات مضاعفة المادة الجينية بمعنى التكرار معقدة وتتطلب إنفاقًا كبيرًا للمواد والطاقة داخل الخلية. لهذا السبب ، يمكن أن تحدث أخطاء النسخ المتماثل العفوي بسهولة. إذا تغيرت المادة الجينية تلقائيًا أو تم إحداثها من الخارج ، فإننا نتحدث عن الطفرات.
يمكن أن تؤدي أخطاء النسخ المتماثل إلى قواعد مفقودة أو مرتبطة بقواعد معدلة أو تكون نتيجة اقتران أساسي غير صحيح. بالإضافة إلى ذلك ، يمكن أن يؤدي حذف وإدخال نيوكليوتيدات مفردة أو متعددة داخل خيطي DNA إلى حدوث أخطاء في النسخ المتماثل. وينطبق الشيء نفسه على ثنائيات البيريميدين ، وفواصل الجدائل وأخطاء الربط المتبادل في خيوط الحمض النووي.
تتوفر آليات إصلاح منفصلة في حالة حدوث خطأ في النسخ المتماثل. يتم تصحيح العديد من الأخطاء المذكورة قدر الإمكان بواسطة بوليميراز الحمض النووي. دقة النسخ المتماثل عالية نسبيًا. معدل الخطأ هو خطأ واحد فقط لكل نوكليوتيد ، والذي يمكن أن يعزى إلى أنظمة تحكم مختلفة.
على سبيل المثال ، تُعرف آلية التحكم في الخلايا حقيقية النواة باسم تسوس الرنا المرسال غير المعقول ، والذي يمكنه التعرف على أكواد الإيقاف غير المرغوب فيها داخل الرنا المرسال وبالتالي منع البروتينات المختصرة من إيجاد التعبير.
ترجع أكواد الإيقاف المبكرة في الرنا المرسال إلى طفرات جينية. يمكن أن يؤدي ما يسمى بالطفرات غير المنطقية أو التضفير الخاطئ والبديل إلى تقصير البروتينات التي تتأثر بفقد وظيفي. لا تستطيع آليات التحكم دائمًا تصحيح الأخطاء.
هناك ثلاثة أشكال مختلفة من اضطراب وراثي وراثي جسمي متنحي β- ثلاسيميا: الأول هو الثلاسيميا متماثلة اللواقح ، وهو مرض خطير يمكن إرجاعه إلى الطفرة غير المنطقية. تعتبر الثلاسيميا متغايرة الزيجوت مرضًا أكثر اعتدالًا حيث توجد طفرات لا معنى لها في نسخة واحدة فقط من جين β-globin. من خلال آلية اضمحلال الرنا المرسال الذي لا معنى له ، يمكن أن يتحلل الرنا المرسال للجين المعيب إلى الحد الذي يتم فيه التعبير عن الجينات السليمة فقط.
في الثلاسيميا متغايرة الزيجوت وبالتالي الشكل المعتدل للمرض ، تكون الطفرة غير المنطقية في إكسون الرنا المرسال الأخير ، بحيث لا يتم تنشيط آليات التحكم. لهذا السبب ، بالإضافة إلى β-globin الصحي ، يتم أيضًا تكوين β-globin المختصر.تموت كريات الدم الحمراء مع-globin المعيب.
مثال آخر على فشل آلية التحكم هو الحثل العضلي الدوشيني ، والذي يرجع أيضًا إلى طفرة غير منطقية في الرنا المرسال. في هذه الحالة ، تقوم آلية التحكم بتفكيك الرنا المرسال ، ولكنها تتسبب بالتالي في خسارة كاملة لما يسمى بروتين ديستروفين.





.jpg)